REKLAMA
Dziennik Ustaw - rok 2017 poz. 1470
ROZPORZĄDZENIE
MINISTRA ROLNICTWA I ROZWOJU WSI1)
z dnia 27 lipca 2017 r.
zmieniające rozporządzenie w sprawie stawek dotacji przedmiotowych dla różnych podmiotów wykonujących zadania na rzecz rolnictwa
Na podstawie art. 130 ust. 5 ustawy z dnia 27 sierpnia 2009 r. o finansach publicznych (Dz. U. z 2016 r. poz. 1870, z późn. zm.2)) zarządza się, co następuje:
§ 1. [Rozporządzenie w sprawie stawek dotacji przedmiotowych dla różnych podmiotów wykonujących zadania na rzecz rolnictwa]
1) w § 5 w ust. 3 pkt 2 otrzymuje brzmienie:
„2) równej 120% ceny targowiskowej ziemniaka w województwie, w którym zostały wyprodukowane te bulwy, ustalonej na podstawie ostatnich danych uzyskanych w badaniach statystycznych statystyki publicznej, dostępnych w dniu sprzedaży tych bulw – jeżeli bulwy ziemniaka zostały wyprodukowane poza umową kontraktacji albo jeżeli umowa kontraktacji nie zawiera ceny sprzedaży tych bulw”;
2) w § 5a w ust. 3 pkt 2 otrzymuje brzmienie:
„2) równej 120% ceny targowiskowej ziemniaka w województwie, w którym zostały wyprodukowane te bulwy, ustalonej na podstawie ostatnich danych uzyskanych w badaniach statystycznych statystyki publicznej, dostępnych w dniu sprzedaży tych bulw – jeżeli bulwy ziemniaka zostały wyprodukowane poza umową kontraktacji albo jeżeli umowa kontraktacji nie zawiera ceny sprzedaży tych bulw”;
3) w § 6 w ust. 3 pkt 2 otrzymuje brzmienie:
„2) równej 120% ceny targowiskowej ziemniaka w województwie, w którym zostały wyprodukowane te bulwy, ustalonej na podstawie ostatnich danych uzyskanych w badaniach statystycznych statystyki publicznej, dostępnych w dniu zniszczenia tych bulw przez ich spalenie w spalarni odpadów lub umieszczenie na składowisku odpadów – jeżeli bulwy ziemniaka zostały wyprodukowane poza umową kontraktacji albo jeżeli umowa kontraktacji nie zawiera ceny sprzedaży tych bulw.”;
4) w § 6a w ust. 3 w:
a) pkt 1 lit. b otrzymuje brzmienie:
„b) 70% kwoty, którą wnioskodawca mógłby uzyskać ze sprzedaży bulw ziemniaka przekazanych jednostce organizacyjnej lub osobie fizycznej, o których mowa w ust. 1, po cenie targowiskowej ziemniaka w województwie, w którym zostały wyprodukowane te bulwy, ustalonej na podstawie ostatnich danych uzyskanych w badaniach statystycznych statystyki publicznej, dostępnych w dniu przekazania tych bulw – jeżeli bulwy ziemniaka zostały wyprodukowane poza umową kontraktacji albo jeżeli umowa kontraktacji nie zawiera ceny sprzedaży tych bulw;”,
b) pkt 2 lit. b otrzymuje brzmienie:
„b) 50% ceny targowiskowej ziemniaka w województwie, w którym zostały wyprodukowane te bulwy, ustalonej na podstawie ostatnich danych uzyskanych w badaniach statystycznych statystyki publicznej, dostępnych w dniu przekazania tych bulw – jeżeli bulwy ziemniaka zostały wyprodukowane poza umową kontraktacji albo jeżeli umowa kontraktacji nie zawiera ceny sprzedaży tych bulw.”;
5) w § 6b w ust. 3 w:
a) pkt 1 lit. b otrzymuje brzmienie:
„b) targowiskowej ziemniaka w województwie, w którym zostały wyprodukowane te bulwy, ustalonej na podstawie ostatnich danych uzyskanych w badaniach statystycznych statystyki publicznej, dostępnych w dniu sprzedaży tych bulw – jeżeli bulwy ziemniaka zostały wyprodukowane poza umową kontraktacji albo jeżeli umowa kontraktacji nie zawiera ceny sprzedaży tych bulw”,
b) pkt 2 lit. b otrzymuje brzmienie:
„b) targowiskowej ziemniaka w województwie, w którym zostały wyprodukowane te bulwy, ustalonej na podstawie ostatnich danych uzyskanych w badaniach statystycznych statystyki publicznej, dostępnych w dniu sprzedaży tych bulw – jeżeli bulwy ziemniaka zostały wyprodukowane poza umową kontraktacji albo jeżeli umowa kontraktacji nie zawiera ceny sprzedaży tych bulw”;
6) w § 6c w ust. 3 w:
a) pkt 1 lit. b otrzymuje brzmienie:
„b) 70% kwoty, którą wnioskodawca mógłby uzyskać ze sprzedaży unieszkodliwionych bulw ziemniaka po cenie targowiskowej ziemniaka w województwie, w którym zostały wyprodukowane te bulwy, ustalonej na podstawie ostatnich danych uzyskanych w badaniach statystycznych statystyki publicznej, dostępnych w dniu unieszkodliwienia tych bulw – jeżeli bulwy ziemniaka zostały wyprodukowane poza umową kontraktacji albo jeżeli umowa kontraktacji nie zawiera ceny sprzedaży tych bulw;”,
b) pkt 2 lit. b otrzymuje brzmienie:
„b) 50% ceny targowiskowej ziemniaka w województwie, w którym zostały wyprodukowane te bulwy, ustalonej na podstawie ostatnich danych uzyskanych w badaniach statystycznych statystyki publicznej, dostępnych w dniu unieszkodliwienia tych bulw – jeżeli bulwy ziemniaka zostały wyprodukowane poza umową kontraktacji albo jeżeli umowa kontraktacji nie zawiera ceny sprzedaży tych bulw.”;
7) w § 8:
a) ust. 4 otrzymuje brzmienie:
„4. Minister właściwy do spraw rolnictwa ogłasza corocznie do dnia 30 września, w dzienniku urzędowym tego ministra, listy, o których mowa w ust. 3, na rok następny oraz zamieszcza je na stronie internetowej administrowanej przez urząd obsługujący tego ministra.”,
b) ust. 6 i 7 otrzymują brzmienie:
„6. Wniosek o udzielenie dotacji składa się do ministra właściwego do spraw rolnictwa w terminie do dnia 31 grudnia roku poprzedzającego rok, w którym udziela się dotacji, zgodnie ze wzorem nr 10 określonym w załączniku nr 2 do rozporządzenia.
7. Minister właściwy do spraw rolnictwa corocznie, do dnia 30 kwietnia, wydaje decyzje w sprawie udzielenia dotacji, z tym że w przypadku gdy organizacja badawcza złożyła więcej niż jeden wniosek o udzielenie dotacji, wydaje jedną decyzję.”;
8) w załączniku nr 2 do rozporządzenia:
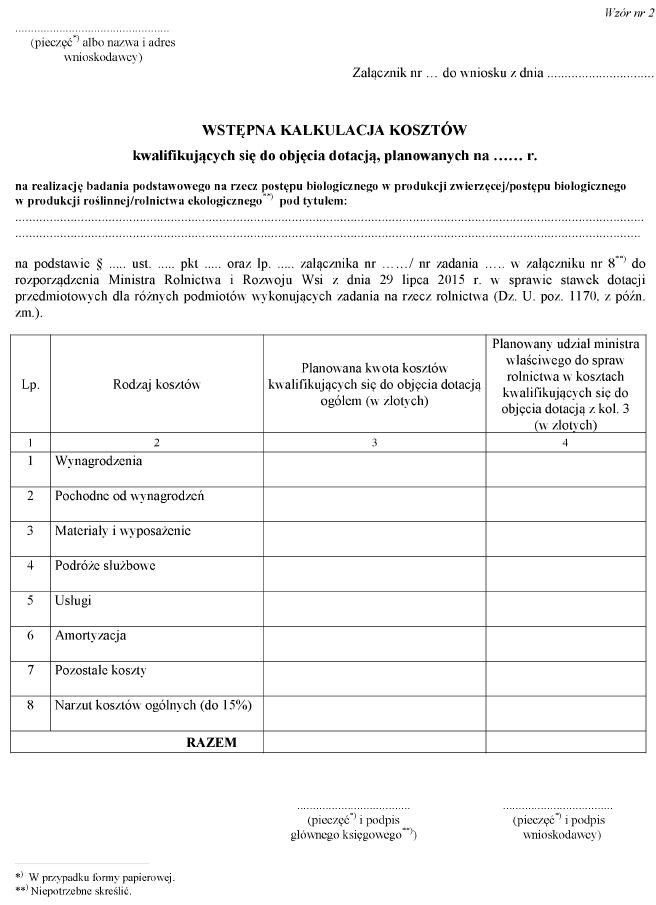
a) wzór nr 2 otrzymuje brzmienie określone w załączniku nr 1 do niniejszego rozporządzenia,
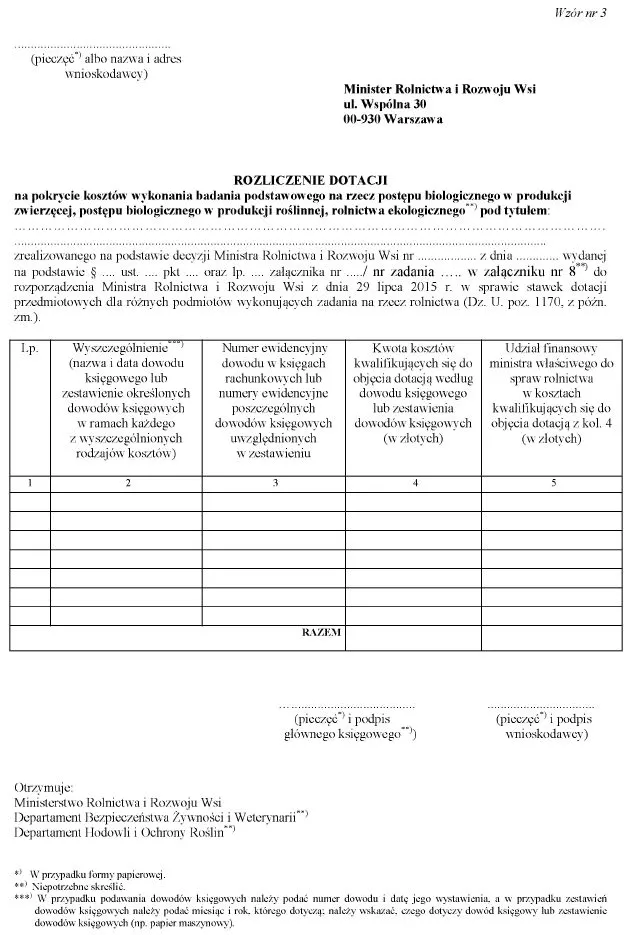
b) wzór nr 3 otrzymuje brzmienie określone w załączniku nr 2 do niniejszego rozporządzenia;
9) załącznik nr 8 do rozporządzenia otrzymuje brzmienie określone w załączniku nr 3 do niniejszego rozporządzenia.
§ 2. [Przepisy dotychczasowe]
§ 3. [Termin ogłoszenia list w 2017 r.]
§ 4. [Przepisy przejściowe]
2. W 2017 r. w odniesieniu do zadań określonych w lp. 3, 6, 10, 56, 70, 79–81, 84, 89 i 95 w tabeli w załączniku nr 8 do rozporządzenia zmienianego w § 1:
1) wniosek o udzielenie dotacji, o których mowa w § 9 ust. 2 rozporządzenia zmienianego w § 1, składa się w terminie 30 dni od dnia wejścia w życie niniejszego rozporządzenia;
2) do rozpatrzenia wniosków stosuje się przepisy dotychczasowe.
3. W przypadku wniosków, o których mowa w ust. 1 i 2, wypłaty, o której mowa w § 9 ust. 7 pkt 1 rozporządzenia zmienianego w § 1, dokonuje się w terminie 90 dni od dnia wejścia w życie niniejszego rozporządzenia.
§ 5. [Wejście w życie]
Minister Rolnictwa i Rozwoju Wsi: K. Jurgiel
1) Minister Rolnictwa i Rozwoju Wsi kieruje działami administracji rządowej - rolnictwo i rozwój wsi, na podstawie § 1 ust. 2 pkt 1 i 2 rozporządzenia Prezesa Rady Ministrów z dnia 17 listopada 2015 r. w sprawie szczegółowego zakresu działania Ministra Rolnictwa i Rozwoju Wsi (Dz. U. poz. 1906).
2) Zmiany tekstu jednolitego wymienionej ustawy zostały ogłoszone w Dz. U. z 2016 r. poz. 1948, 1984 i 2260 oraz z 2017 r. poz. 60, 191, 659, 933, 935 i 1089.
Załączniki do rozporządzenia Ministra Rolnictwa i Rozwoju Wsi
z dnia 27 lipca 2017 r. (poz. 1470)
Załącznik nr 1
WZÓR – WSTĘPNA KALKULACJA KOSZTÓW
Załącznik nr 2
WZÓR – ROZLICZENIE DOTACJI
Załącznik nr 3
STAWKI DOTACJI NA POKRYCIE KOSZTÓW BADAŃ PODSTAWOWYCH NA RZECZ POSTĘPU BIOLOGICZNEGO W PRODUKCJI ROŚLINNEJ
| Lp. | Nr zadania | Zadanie | Stawka |
| 1 | 2 | 3 | 4 |
| 1 | 2 | Wykorzystanie markerów molekularnych i fenotypowych do identyfikacji genów odporności pszenicy na łamliwość źdźbła powodowaną przez Oculimacula yallundae i Oculimacula acuformis | 108 000 |
| 2 | 3 | Badania nad wpływem translokacji 1B/1R na efektywność uzyskiwania linii DH pszenicy oraz ich wartość technologiczną | 179 100 |
| 3 | 4 | Mapowanie asocjacyjne genów odporności na rdzę brunatną (Puccinia triticina) i septoriozę paskowaną liści (Septoria tritici) w pszenicy | 147 600 |
| 4 | 5 | Identyfikacja zmienności genetycznej pszenicy korelującej z potencjałem plonotwórczym i wybranymi cechami systemu korzeniowego | 135 000 |
| 5 | 6 | Poszukiwanie oraz wykorzystanie markerów fenotypowych, metabolicznych i molekularnych do badania typów odporności na fuzariozę kłosów u form pszenicy o zróżnicowanej podatności | 397 800 |
| 6 | 8 | Tolerancja na stresy abiotyczne – genotypowanie pszenicy w oparciu o strategię genów kandydujących | 135 000 |
| 7 | 9 | Efektywność piramidowania genów odporności na mączniaka prawdziwego (Blumeria graminis f. sp. tritici) i rdzę brunatną (Puccinia triticina) w pszenicy ozimej | 139 500 |
| 8 | 10 | Toksyny białkowe Parastagonospora nodorum i ich związek z patogenicznością oraz odpornością pszenżyta i pszenicy na septoriozę liści i plew | 197 100 |
| 9 | 11 | Określenie czynników decydujących o zimowaniu pszenicy ozimej i pszenżyta ozimego w warunkach polskich | 135 000 |
| 10 | 12 | Analiza zmienności somaklonalnej indukowanej w kulturach in vitro u roślin zbożowych | 72 000 |
| 11 | 13 | Opracowanie i wykorzystanie metod biotechnologicznych do skrócenia cyklu hodowlanego pszenżyta oraz do poprawy efektywności selekcji – miejscowo-specyficzna mutageneza z wykorzystaniem miejscowo-specyficznych nukleaz | 157 800 |
| 12 | 14 | Badanie typów odporności pszenżyta ozimego na fuzariozę kłosów za pomocą markerów fenotypowych i metabolicznych | 166 500 |
| 13 | 15 | Poszukiwanie markerów molekularnych genów utrzymania sterylności pyłku u pszenżyta z cms Tt | 139 500 |
| 14 | 17 | Wytwarzanie nowych źródeł genetycznych pszenżyta w oparciu o krzyżowanie oddalone | 72 000 |
| 15 | 19 | Badania nad zwiększeniem odporności żyta na sporysz i na fuzariozę kłosów przez poznanie interakcji pasożyt – żywiciel – środowisko z wykorzystaniem genetycznych źródeł odporności na Claviceps purpurea i grzyby rodzaju Fusarium | 93 400 |
| 16 | 20 | Poszukiwanie wspólnych mechanizmów dziedziczenia płodności roślin z cytoplazmą CMS-C oraz z cytoplazmą CMS-Pampa | 171 000 |
| 17 | 21 | Poszukiwanie markerów molekularnych genów przywracania płodności pyłku u żyta (Secale cereale L.) z CMS-Pampa | 139 500 |
| 18 | 22 | Poszukiwanie źródeł genetycznej odporności na mączniaka i rdzę w kolekcji linii, rodów i odmian żyta | 90 000 |
| 19 | 23 | Analiza zmienności epigenetycznej indukowanej stresem suszy oraz ocena jej stabilności transgeneracyjnej w aspekcie tolerancji jęczmienia na stres niedoboru wody | 139 500 |
| 20 | 24 | Badania nad wpływem brasinosteroidów na tolerancję roślin jęczmienia na stres niedoboru wody | 84 600 |
| 21 | 25 | Molekularne podstawy zjawiska albinizmu w kulturach izolowanych mikrospor jęczmienia | 192 500 |
| 22 | 26 | Identyfikacja czynników determinujących odporność jęczmienia ozimego (Hordeum vulgare L.) na suszę i mróz | 113 400 |
| 23 | 27 | Współdziałanie odporności na mączniaka (Blumeria graminis f.sp. hordei) warunkowanej genem mlo z wartością cech gospodarczych jęczmienia ozimego | 126 000 |
| 24 | 28 | Badania zdrowotności owsa (Avena sativa L.) z uwzględnieniem biologii i szkodliwości wybranych patogenów dla tego zboża | 81 000 |
| 25 | 29 | Poszukiwanie źródeł odporności owsa (Avena sativa L.) na nowy patogeniczny i mykotoksynotwórczy gatunek – Fusarium langsethiae | 34 200 |
| 26 | 30 | Mapowanie sprzężeniowe i asocjacyjne owsa zwyczajnego | 180 000 |
| 27 | 31 | Piramidyzacja genów odporności na rdzę koronową w genomie owsa oraz identyfikacja i lokalizacja markerów DNA dla tych genów | 72 000 |
| 28 | 32 | Badanie składników determinujących wartość odżywczą i funkcjonalną owsa oraz ich relacji w ziarnie obłuszczonym i oplewionym | 87 300 |
| 29 | 33 | Poszukiwanie form kukurydzy o wysokiej odporności na fuzariozę kolb i zgorzel podstawy łodygi powodowane przez grzyby z rodzaju Fusarium spp. | 117 000 |
| 30 | 34 | Określenie zróżnicowania genetycznego linii wsobnych kukurydzy za pomocą markerów molekularnych | 76 500 |
| 31 | 35 | Identyfikacja genów związanych z ekspresją zimotrwałości i tolerancji suszy u form introgresywnych Lolium multiflorum/Festuca arundinacea | 135 000 |
| 32 | 36 | Gromadzenie i ocena kolekcji ekotypów traw wieloletnich z uwzględnieniem cech warunkujących ich wykorzystanie na cele alternatywne | 144 000 |
| 33 | 38 | Badanie cech warunkujących zawiązywanie nasion, ich jakość oraz plon w wybranych gatunkach traw wieloletnich | 135 900 |
| 34 | 39 | Cecha wczesności kwitnienia u łubinu białego i łubinu żółtego – podstawy genetyczne i molekularne | 117 000 |
| 35 | 40 | Identyfikacja rejonów w genomie grochu, warunkujących wybrane parametry sprawności fizjologicznej, jako istotnego elementu odporności na stresy abiotyczne | 144 000 |
| 36 | 41 | Identyfikacja i sposób dziedziczenia genów warunkujących odporność na choroby grzybowe i niską zawartość alkaloidów w doskonaleniu wartości użytkowej łubinów, ze szczególnym uwzględnieniem łubinu żółtego | 164 700 |
| 37 | 42 | Analiza zmienności genetycznej i piramidyzacja genów warunkujących cechy użytkowe łubinu białego | 67 500 |
| 38 | 43 | Analiza bioróżnorodności zasobów genowych soi przydatnej do hodowli w warunkach klimatycznych Polski i opracowanie metodyki krzyżowania międzygatunkowego Glycine max x Glycine soja | 72 000 |
| 39 | 44 | Identyfikacja i zastosowanie fenotypowych i molekularnych markerów tolerancji na stres suszy u buraka cukrowego | 160 500 |
| 40 | 45 | Opracowanie metod globalnej analizy polimorfizmów w genomie buraka cukrowego | 207 000 |
| 41 | 46 | Badania nad mechanizmami warunkującymi proces embriogenezy gametycznej u buraka cukrowego | 126 000 |
| 42 | 48 | Badanie genomu rzepaku ozimego przy wykorzystaniu markerów molekularnych | 297 000 |
| 43 | 49 | Badanie bioróżnorodności gatunków z plemienia Brassiceae w celu otrzymania form rzepaku ulepszonych pod względem odporności na patogeny | 99 000 |
| 44 | 50 | Zastosowanie konwencjonalnych i molekularnych narzędzi fitopatologicznych w poszukiwaniu źródeł odporności na kiłę kapusty oraz charakterystyka aktualnej populacji patogenu w Polsce | 193 500 |
| 45 | 51 | Wprowadzanie nowych alleli z pul genowych różnych gatunków z rodzaju Brassica do bazy genowej rzepaku ozimego | 117 000 |
| 46 | 53 | Wykorzystanie nowej puli genowej dla uzyskania form rzepaku ozimego o zmienionych cechach jakościowych | 225 000 |
| 47 | 54 | Introdukcja genów odporności na choroby i owady oraz męskiej sterylności z pokrewnych gatunków rodzaju Brassica do rzepaku (Brassica napus L.) | 144 000 |
| 48 | 55 | Opracowanie modeli kalibracyjnych dla spektrometru NIRS o zakresie widma 400–2500 nm dla oznaczania glukozynolanów, białka, NDF, ADF oraz steroli i badania zmienności tych związków w roślinach oleistych | 130 500 |
| 49 | 56 | Badania ekspresji i genetyczna charakterystyka odporności na bakterie Dickeya solani w wyróżnionych źródłach odporności w ziemniaku | 301 500 |
| 50 | 57 | Badania nad opracowaniem metod selektywnej izolacji oraz czułej identyfikacji bakterii Clavibacter michiganensis ssp. sepedonicus w trudnych diagnostycznie próbach środowiskowych | 360 000 |
| 51 | 58 | Opracowanie czułych metod wykrywania najważniejszych wirusów ziemniaka | 321 300 |
| 52 | 59 | Badania tolerancji odmian ziemniaka na stresy abiotyczne w świetle postępujących zmian klimatycznych | 180 000 |
| 53 | 60 | Wyróżnianie form ziemniaka o złożonej odporności na mątwiki atakujące ziemniak przy wykorzystaniu metod konwencjonalnych i molekularnych. Charakterystyka nowego źródła odporności na Globodera pallida znalezionego w Solarium gourlayi | 138 600 |
| 54 | 61 | Wyróżnianie i charakterystyka tetraploidalnych form ziemniaka odpornych na wirusy M i S ziemniaka z wykorzystaniem selekcji metodami konwencjonalnymi i markerami molekularnymi | 128 700 |
| 55 | 62 | Analiza interakcji genotypowo-środowiskowej w odniesieniu do wybranych cech użytkowych ziemniaka jadalnego w różnych systemach uprawy | 154 800 |
| 56 | 63 | Eliminacja patogenów niekwarantannowych (bakterie endogenne i wirusy) oraz kontrola zdrowotności roślin ziemniaka w banku in vitro | 108 000 |
| 57 | 64 | Wykorzystanie metod biotechnologicznych do poszerzenia zmienności genetycznej warzyw kapustnych | 198 000 |
| 58 | 66 | Badania nad opracowaniem molekularnej metody identyfikacji genów warunkujących ważne cechy użytkowe pomidora | 189 000 |
| 59 | 67 | Otrzymanie nowej zmienności genetycznej warzyw kapustowatych przy wykorzystaniu krzyżowań oddalonych w rodzaju Brassica | 180 000 |
| 60 | 68 | Analiza czynników genetycznych związanych z przywracaniem płodności roślin buraka ćwikłowego | 99 000 |
| 61 | 69 | Opracowanie i wykorzystanie wysoko wydajnych technik selekcji genomowej w doskonaleniu warzyw | 198 000 |
| 62 | 70 | Indukowanie zmienności genetycznej jabłoni na drodze poliploidyzacji in vitro oraz ocena fenotypowa i genetyczna uzyskanych poliploidów w odniesieniu do diploidalnych form wyjściowych | 92 500 |
| 63 | 71 | Analiza genetyczna i molekularna wybranych genotypów jabłoni (Malus domestica) dla skrócenia okresu juwenilnego i poprawy jakości owoców | 243 000 |
| 64 | 72 | Ocena potencjału genetycznego wybranych genotypów borówki wysokiej (Vaccinium corymbosum L.) w oparciu o czynnikowy układ krzyżowań | 157 500 |
| 65 | 73 | Poszukiwanie regionów DNA sprzężonych z tolerancją wegetatywnych podkładek jabłoni na niskie temperatury poprzez analizę transkryptomu i ocenę stopnia polimorfizmu genów kandydujących | 171 000 |
| 66 | 74 | Badania nad saturacją mapy genetycznej Elsanta x Senga Sengana pod kątem lokalizacji genów sprzężonych z ważnymi cechami użytkowymi truskawki (Fragaria x ananassa) | 171 000 |
| 67 | 75 | Badania nad możliwością poszerzenia zmienności genetycznej maliny właściwej (Rubus idaeus) pod względem różnej pory dojrzewania i jakości owoców | 157 500 |
| 68 | 76 | Badania nad możliwością zwiększenia zawartości składników bioaktywnych w owocach truskawki na drodze hybrydyzacji wewnątrz- i międzygatunkowej w obrębie rodzaju Fragaria | 171 000 |
| 69 | 77 | Hybrydyzacja oddalona gatunków Prunus cerasifera (ałycza), Prunus armeniaca (morela), Prunus salicina (śliwa japońska), Prunus domestica (śliwa domowa) w celu zwiększenia bioróżnorodności genetycznej w obrębie rodzaju Prunus | 171 000 |
| 70 | 78 | Charakterystyka markerów molekularnych, sprzężonych z odpornością na wielkopąkowca porzeczkowego (Cecidophyopsis ribis) | 128 300 |
| 71 | 79 | Analiza czynników warunkujących organogenezę agrestu (Ribes grossularia L.) w kulturach in vitro i in vivo oraz ocena genetyczna i fenotypowa otrzymanego materiału | 117 000 |
| 72 | 80 | Alternatywne czynniki redukujące zjawisko albinizmu roślin regenerowanych z androgenicznych kultur pszenicy i pszenżyta | 168 800 |
| 73 | 81 | Opracowanie i wykorzystanie metod biotechnologicznych skracających cykl hodowlany i zwiększających efektywność selekcji genotypów ozimej pszenicy i ozimego pszenżyta o podwyższonej odporności i tolerancji na septoriozę liści i plew [czynnik sprawczy: Parastagonospora nodorum (Berk.), (Quaedvlieg, Verkley & Crous.)]. | 202 500 |
| 74 | 82 | Identyfikacja regionów genomu oraz markerów DNA związanych z heterozją w heksaploidalnym pszenżycie ozimym | 144 900 |
| 75 | 83 | Genetyczne podłoże męskiej sterylności pszenżyta z różnymi cytoplazmami oraz możliwość wykorzystania badanych cytoplazm do tworzenia systemów CMS u pszenicy | 180 000 |
| 76 | 84 | Identyfikacja czynników determinujących efektywność otrzymywania podwojonych haploidów żyta (Secale cereale L.) metodami androgenezy i krzyżowań oddalonych | 158 400 |
| 77 | 85 | Badanie reakcji mikrospor żyta na stres i warunki kultury in vitro | 135 000 |
| 78 | 86 | Haploidyzacja żyta – diagnostyka molekularna oraz wpływ nanomolekuł na wspomaganie indukcji i regeneracji roślin w warunkach in vitro | 81 000 |
| 79 | 87 | Badania wewnętrznej struktury genetycznej odmian żyta oraz dziedzicznego podłoża efektu heterozji | 180 000 |
| 80 | 88 | Efekty plejotropowe genów Ppd-H1 i Ppd-H2 a podatność roślin jęczmienia jarego na fuzariozę kłosów i akumulację mikotoksyn | 171 000 |
| 81 | 89 | Molekularna charakterystyka wpływu elementów mobilnych na zmienność genetyczną w zbożowych kulturach in vitro | 81 000 |
| 82 | 91 | Identyfikacja i lokalizacja markerów DNA dla wybranych genów odporności na mączniaka prawdziwego w owsie zwyczajnym oraz piramidyzacja efektywnych genów odporności w genomie owsa | 81 000 |
| 83 | 92 | Fusarium temperatum – znaczenie i szkodliwość w uprawie kukurydzy, poszukiwanie i charakterystyka źródeł odporności | 216 000 |
| 84 | 93 | Fenotypowanie i genotypowanie łubinu wąskolistnego pod względem wybranych cech morfologicznych, plonotwórczych i parametrów technologicznych nasion | 66 700 |
| 85 | 94 | Badanie czynników determinujących niską strawność białka śruty uzyskanej z nasion rzepaku ozimego | 132 900 |
| 86 | 95 | Badania nad regulatorową funkcją cząsteczek miRNA w przebiegu infekcji wirusem ziemniaka PVY | 150 000 |
| 87 | 96 | Badania cytologicznych i biochemicznych mechanizmów odporności roślin w patosystemach pomidor – Phytophtora infestans oraz ogórek – Pseudoperonospora cubensis | 124 300 |
| 88 | 97 | Identyfikacja genów odpowiedzialnych za przywracanie płodności i samozgodność u wybranych roślin warzywnych | 135 000 |
| 89 | 98 | Analiza wpływu hipoksji na zwiększenie tolerancji na stresy u pomidora (Lycopersicon esculentum L.) i ogórka (Cucumis sativus L.) | 199 800 |
| 90 | 99 | Badanie molekularnego mechanizmu odporności na kiłę kapusty (Plasmodiochora brassicae) u roślin z rodzaju Brassica | 162 000 |
| 91 | 100 | Doskonalenie ogórka (Cucumis sativus L.) pod względem odporności na kanciastą plamistość | 156 200 |
| 92 | 101 | Transfer cytoplazmatycznej męskiej sterylności poprzez somatyczną hybrydyzację u marchwi | 126 000 |
| 93 | 102 | Opracowanie genetycznych, fizjologicznych i biochemicznych podstaw tolerancji ogórka na stres niedoboru wody | 144 000 |
| 94 | 103 | Fizjologiczne i genetyczne markery odporności pszenicy jarej na suszę w aspekcie interakcji zachodzących w ryzosferze | 144 700 |
| 95 | 104 | Analiza genetycznych i biochemicznych podstaw tolerancji cebuli (Allium cepa L.) na stres niedoboru wody | 171 000 |
| 96 | 105 | Identyfikacja układów allelicznych genów fotoneutralności i wczesności oraz opracowanie metodyki otrzymywania roślin homozygotycznych u soi | 253 800 |
| 97 | 106 | Wpływ stresu niedoboru wody na rozwój i architekturę systemu korzeniowego u jęczmienia (Hordeum vulgare L.) | 278 600 |
| 98 | 107 | Występowanie dotychczas niemonitorowanych wirusów (HpLV, ArMV) i wiroidów (HpSVd, AFCVd, CBCVd) na plantacjach produkcyjnych chmielu w Polsce | 298 800 |
| 99 | 108 | Badania nad gametyczną embriogenezą u Lupinus angustifolius L. – indukcja haploidów i analiza genetycznego podłoża tego procesu | 120 000 |
| 100 | 109 | Selekcja genomowa pszenicy ozimej | 337 500 |
- Data ogłoszenia: 2017-07-31
- Data wejścia w życie: 2017-08-01
- Data obowiązywania: 2017-08-01

REKLAMA
Dziennik Ustaw
REKLAMA
REKLAMA